Die Pflanzenschrift und ihre Offenbarungen
von Jagadis Chunder Bose
XXIV. KAPITEL
BESTIMMUNG DER LAGE DES NERVEN
Nachdem es sichergestellt war, daß es in der Pflanze eine nervöse Reizleitung gibt, bestand die nächste Aufgabe darin, festzustellen, welches das leitende Gewebe ist und wo es liegt. Der Durchgang eines Reizimpulses durch einen Nerven bewirkt keine sichtbare Veränderung; sondern wir können den Durchgang des Reizes nur durch den negativ elektrischen Ausschlag nachweisen, von dem er begleitet ist. Der Nerv ist in nicht leitendes Gewebe eingebettet und läßt sich mit einem elektrischen Kabel vergleichen, das von einer nichtleitenden Guttaperchaschicht umhüllt ist. In einem Kabel kann ein einzelner Leitungsdraht liegen oder zwei solche. Wir können Telegramme im Kabel auffangen, indem wir eine Metallnadel in dasselbe einführen, die in geeigneter Weise mit einem Galvanometer verbunden ist. Wir werden solange keine Nachrichten auffangen können, bis die Nadel eben das leitende Metall berührt. Aus der Tiefe, bis zu der die Nadel eindringen muß, erfahren wir, wie tief der leitende Metalldraht gelegen ist.
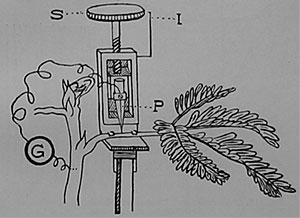 |
| Fig. 111. Elektrische Probe zur Bestimmung der Lage des Nervengewebes im Hauptblattstiel von Mimosa. P die Sonde in Verbindung mit dem Galvanometer G, S Schraubenkopf, durch dessen Drehung die Sonde in das Gewebe eingeführt wird, I Skala, an der die Tiefe der Einstoßung gemessen wird. |
Nach diesem Prinzip vorgehend, gelang es mir, die Lage des Nerven, der in nicht leitendes Gewebe eingebettet ist, zu bestimmen (Fig. 111); ich habe dabei die elektrische Probe angewandt, die schon in einem früheren Kapitel beschrieben worden ist. Das äußere Ende eines Blättchenstieles von Mimosa wird periodisch gereizt, während die Sonde schrittweise, z. B. in Schritten von je 0,05 mm in den Blattstiel eingeführt wird. Am Beginn läßt sich kein elektrischer Ausschlag, wie er für den nervösen Impuls kennzeichnend ist, nachweisen; wir müssen die Sonde tiefer führen. Jetzt beginnen die ersten Meldungen; und bei einer gewissen Tiefe werden die elektrischen Ausschläge am stärksten. Wir merken uns diese Tiefe an. Wird die Sonde tiefer eingestoßen, so verschwindet die elektrische Reaktion wieder; die Sonde hat das leitende Gewebe durchdrungen und ist in nicht leitende Gewebsschichten eingetreten. Wird sie noch tiefer geführt, so erhält man wieder Meldungen, jenseits dieser Schicht aber gibt es keine Ausschläge mehr. Auf diese Weise ist es möglich, die Lage des leitenden Gewebes bis auf einen Hundertstel-Zoll genau zu bestimmen. Die Beobachtungen ergeben, daß die Erregungsleitung auf ein bestimmtes Gewebe sich beschränkt, welches daher als Nerv bezeichnet werden darf.
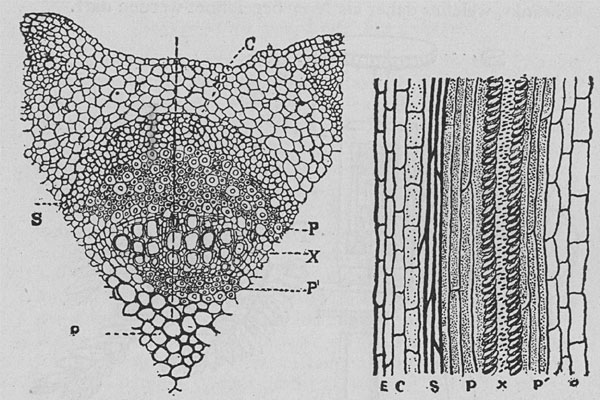 |
| Fig. 112. Querschnitt und Längsschnitt durch ein einzelnes Gefäßbündel von Mimosa. Figur links: Querschnitt. Die punktierte senkrechte Linie bezeichnet die Einstichlinie der elektrischen Sonde. C Rinde, S Sklerenchyn, P äußeres Phloem, X Xylem, P' inneres Phloem, O Mark. Figur rechts: Längsschnitt durch das Bündel. Man beachte die langgestreckten zylindrischen Zellen im äußeren und im inneren Phloem. (Der Schnitt hat das Bündel nur seitlich, nicht durch die Mitte getroffen.) |
Wir stellen nun einen Längsschnitt durch den Blattstiel her, der die Einstichlinie der Nadel umfaßt, um festzustellen, an welcher Stelle die Sonde die Meldungen aufgefangen hat. Die Epidermis hat solche nicht ergeben; desgleichen war die primäre Rinde offenbar eine nichtleitende Hülle; starke Impulse wurden uns vermeldet, als die Sonde in das Phloem trat. Als sie durch das Xylem oder Holz drang, hörten die Meldungen auf, doch in der nächsten Schicht begannen sie wieder. Die so nachgewiesene zweite leitende Gewebsschicht ist ein zweites, inneres Phloem, dem die Pflanzenphysiologen bisher keine Beachtung geschenkt hatten. Wir haben somit nicht nur eine nervöse Gewebsschicht, sondern zwei solche nachgewiesen (Fig. 112). Die Bedeutung dieser Zweiteilung des Nervensystems in eine äußere und eine innere Lage soll später erläutert werden.
NACHWEIS DER NERVENVERTEILUNG DURCH SELEKTIVE FÄRBUNG
Wenn wir eine Färbungsmethode ausfindig machen können, welche die Nervenstränge hervorhebt, die wir jetzt als die Bahnen der Reizimpulse kennen gelernt haben, dann läßt sich die Verteilung der Nerven in der Pflanze in klarer Weise zur Anschauung bringen. Auf solche Art muß es möglich sein, zwei benachbarte Gewebssysteme von verschiedener Funktion zu unterscheiden oder die gleichartige Funktion zweier räumlich getrennter Gewebe nachzuweisen. Durch Hämatoxilin und Safranin färbte sich das Nervengewebe tief violett und hob sich aufs deutlichste von den anderen Geweben ab. Dieser Befund sichert noch die Ergebnisse der elektrischen Sondierung. Das äußere und innere Phloem waren ganz ähnlich gefärbt, woraus hervorgeht, daß es sich in der Tat um zwei getrennte Nerven handelt. Im Blattstiel finden sich vier solche Doppelstränge, jedes Paar kommt aus einem Blättchenstiel und endigt im Gelenkpolster.
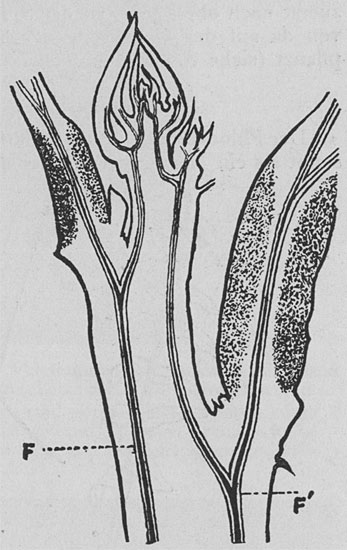 |
| Fig. 113. Zusammenhang des Leitgewebes zwischen Stengel und Blatt. Längsschnitt des Stengels mit einem Blatt auf jeder Seite. Die zwei Gefäßbündel F, F' geben Seitenäste nach den Blättern ab und treffen sich an der Stengelspitze. Das doppelte Phloem ist violett gefärbt und hebt sich vom Grunde ab. Das Gelenkpolster jedes Blattes ist schattiert. |
Im Stengel der Mimosa selbst finden sich zwei gegenüberliegende Hauptgefäßbündelstränge, deren jeder einen doppelten Nervenstrang enthält. Die Hauptsträngegeben seitliche Äste nach den Blättern hin ab, so daß der Zusammenhang des Leitgewebes zwischen Stengel und Blättern gesichert ist. Ein durch Reizung des Stengels bewirkter Impuls kann also in der Richtung zu den Blättern geleitet werden; und ebenso kann ein in den Blättern erzeugter Impuls abwärts zum Stengel wandern, sich hier nach oben und unten nach den anderen Blättern fortpflanzen und deren Fallbewegung bewirken.
 |
| Fig. 114. Blatt eines Farnes; die freigelegten Gefäßbündel N sind in der Figur rechts vergrößert dargestellt. |
Es ist auch bemerkenswert, daß die zwei Hauptstränge der Gefäßbündel an der Spitze des Stammes konvergieren und sich treffen (Fig. 113). Dies erklärt uns, wie es kommt, daß bei mäßig starker Reizung der einen Stengelseite der Impuls zuerst nach oben geht, an der Spitze überspringt und sich von da auf der Gegenseite des Stengels nach unten fortpflanzt.
DER ISOLIERTE PFLANZENNERV
Der Phloemstrang im Gefäßbündel von Mimosa hat sich somit als ein Nerv erwiesen, der die Erregung leitet. Es ist nun unmöglich, den Nerv aus der Mimosa herauszuziehen, ohne ihn in Stücke zu reißen; hingegen gelang es mir, den Nerven aus dem Blattstiel eines Farnes zu isolieren. Die harte Außenschicht des Blattstieles wurde behutsam gebrochen und durch vorsichtiges Auseinanderziehen wurden die Gefäßbündel, die Nervenstränge, freigelegt; sie sind weich, von weißer Farbe und im Aussehen dem tierischen Nerv auffallend ähnlich (Fig. 114).
Wir wollen nun gewisse Versuche, welche die Tierphysiologen üblicherweise am Froschnerven ausführen, am isolierten Nerven der Pflanzen wiederholen. Die Versuche am Froschnerven gelangen mit Hilfe eines Galvanometers zur Ausführung, der die elektrischen Ausschläge, welche den Nervenimpuls begleiten, automatisch verzeichnet. Die elektrischen Registrierkurven des Pflanzennerven unter wechselnden Außenbedingungen fallen in jeder Hinsicht ähnlich aus wie jene des tierischen Nerven. Das folgende Beispiel zeigt dies in schlagender Weise.
 |
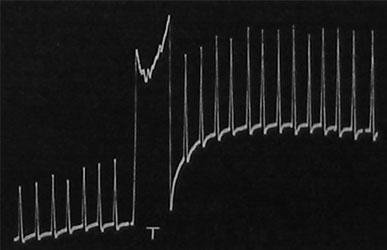 |
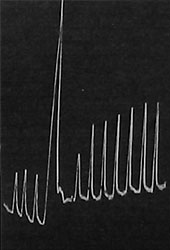
| Fig. 115. Aufzeichnung der Verstärkung der Reaktionsausschläge als Nachwirkung der Wärme-Tetanisation beim Froschnerven. Die drei ersten Reaktionen sind normal; darauf wurde kurze Wärme-Tetanisation angewandt; die nachher durch Reize von der früheren Stärke erhaltenen Ausschläge sind verstärkt. |
| Fig. 116. Photographische Aufzeichnung der Wirkung der Tetanisation T, die zur Verstärkung der normalen Reaktion beim Farn-Nerven führt. |
Es ist eine wohlbekannte Tatsache, daß ein Nerv, der zu lange untätig gewesen ist, mehr oder minder träge wird und daß er durch fortgesetzte Reizung oder durch Tetanisierung zu voller Tätigkeit gebracht werden kann. Die schwachen Reaktionen des trägen Nerven werden nach einer Periode der Tetanisierung bedeutend verstärkt. Dies ist in Fig. 115 dargestellt, in der die drei ersten Ausschläge die Reaktionen des trägen Froschnerven darstellen; nach der Tetanisierung sind die Reaktionen bedeutend stärker als zuvor. Durchaus ähnliche Resultate wurden am Nerven des Farnes erhalten (Fig. 116).
DIE SYNAPTISCHE MEMBRAN
In früherer Zeit wurde die Kontinuität des Protoplasmas als eine wesentliche Voraussetzung der Erregungsleitung in der Pflanze betrachtet. Allein im tierischen Nerv besteht keine protoplasmatische Kontinuität zwischen den End-bäumchen der sich berührenden Neuronen, vielmehr ist die trennende Membran unter dem Namen Synapse bekannt. Diese Membran wirkt ähnlich einem Ventil, indem sie den Impuls im Nerven leichter nach der einen als nach der anderen Richtung fortschreiten läßt. Auch gewisse andere Charakterzüge des nervösen Impulses gehen auf jene Wirkung der Synapsen zurück.
Die eingehende Untersuchung des Nervengewebes der Pflanze zeigt, daß dieses aus verlängert röhrenförmigen Zellen besteht, deren Querwände als synapsenähnliche Membranen wirken.
BAHNUNG
Eine Klappe oder ein Ventil wird offenbar leichter funktionieren, wenn es bei wiederholter Benützung häufig geöffnet wird. Eine Türangel, die von langem Nichtgebrauch rostig ist, wird eines stärkeren Stoßes bedürfen als eine, die häufig benützt wird. Etwas Ähnliches findet man hinsichtlich der Leitung des Nervenimpulses, denn "hier wirkt der Durchgang des Impulses dahin, die Widerstände zu vermindern, so daß ein zweiter einwirkender Reiz die Reaktion leichter hervorruft."5
Diese charakteristische Reaktion ist unter dem Namen "Bahnung" bekannt; durch häufigen Gebrauch wird gleichsam die Bahn wegsam gemacht. Die Eigenschaften des Pflanzennerven erweisen sich jenen des tierischen Nerven sehr ähnlich. Zunächst gibt es, wiewohl der Impuls nach beiden Richtungen wandern kann, doch eine bevorzugte Richtung, in welcher er sich leichter und schneller bewegt, eben infolge der Ventilwirkung der synapsenähnlichen Membranen. Die zentrifugale Erregungsleitung erfolgt leichter als die zentripetale.
Ferner führt vorangegangene Reizung zur Bahnung oder erleichterten Transmission. So war in einem typischen Fall das Reizleitungsvermögen einer Mimosenpflanze so gering, daß der durch einen Normalreiz des Blattstieles im Abstand von 15 mm bewirkte Impuls das Gelenkpolster nicht mehr erreichte. Auf einen viel stärkeren Reiz hin bahnte sich der Impuls trotz des Widerstandes seinen Weg und gelangte rasch bis an das Gelenkpolster. Als nun der Pfad einmal gebahnt war, geschah die Übertragung der folgenden Impulse leicht, und der vorher unwirksame Normalreiz wurde jetzt wirksam.
VERZÄRTELUNG
Im Hinblick auf das Steigen und Sinken der Nervenleistung durch Gebrauch oder Nichtgebrauch sind folgende Beobachtungen von großem Interesse. Eine Pflanze, die man unter einer Glasglocke erzieht und sorgsam vor allen Reizen und Schäden der Außenwelt beschützt, sieht frisch und blühend aus. In Wahrheit ist sie weichlich und dekadent. Anatomisch ist das Nervengewebe vorhanden, doch funktionell ist es infolge mangelnden Gebrauches inaktiv.
Es ist sehr interessant, an einer derartigen Pflanze die Zunahme der Nervenleitfähigkeit unter dem Einfluß äußerer Reize zu beobachten. Zuerst findet keine Transmission statt, nach einiger Zeit beginnt die Übertragung der Erregungsimpulse und fortgesetzte Reizung erhöht das Leitvermögen zu einem Maximum.
Hier liegt ein Fall der Veränderlichkeit des Organismus durch seine Umwelt klar vor uns, durch Summation der Reize kommt das Organ erst zur Ausbildung. Der ungereizte Nerv ist untätig und träge; doch die Reizung verleiht ihm Energie, seine Erregbarkeit und sein Leitvermögen steigt gewaltig an.
DAS BLATT ALS FANGBECKEN FÜR DIE REIZE
Bei der Betrachtung der Reizwirkungen und ihrer Bedeutung für die Erhaltung des Lebensgetriebes in der Pflanze ist folgender Gesichtspunkt von Wichtigkeit. Für die Erhaltung ihrer normalen Funktionstätigkeit müssen die inneren Gewebe in einem günstigen, tonischen Zustand erhalten werden, und zwar durch Reize, die nur von der Außenwelt herkommen können. Unter den äußeren Reizen, die in der Natur auf die Pflanze wirken, ist keiner bedeutsamer als das Licht. Alles ist danach angetan, die Transmission der Lichtreize nach den inneren Geweben zu begünstigen; sie erfolgt längs der Nervenbahnen - längs des Phloems im Stranggewebe. Die Pulsation, die das Saftsteigen unterhält, kommt, wie wir sahen, zum Stillstand, wenn die Pflanze des Lichtes beraubt wird; wird sie dann aber dem Reiz des Lichtes wieder ausgesetzt, so wird der Pulsschlag erneut, der Saft wieder emporgepumpt, und durch die Kanäle, die als Arterien dienen, fließt neues Leben. Das ausgebreitete Blatt, worin die Gefäßbündel als feines Netzwerk verteilt sind, ist nicht bloß ein Gebilde zur Festlegung des Kohlenstoffes aus dem Kohlensäuregas der Luft, sondern auch ein Fangbecken für die Lichtreize, deren Erregungseffekt gesammelt und durch die feineren und die stärkeren Nervenäste nach dem Inneren der Pflanze geleitet wird. Die Verteilung der Gefäßbündel ist dort eine solche, daß kein Teil des lebenden Gewebes zu weit von ihnen abliegt, um die durch die Nervenbahnen zugeleitete Erregung zu empfangen.
So stehen alle Teile der Pflanze durch jenes Nervensystem in inniger und rascher gegenseitiger Verbindung. Nur der Existenz dieses Nervensystems ist es zu danken, daß die Pflanze ein organisches Ganzes, ein Individuum darstellt, worin ein jeder Teil die Reize verspürt, die ein anderer empfängt.
Erläuterung der Fußnoten.
5. Starling, Principles of Human Physiology (1920), S. 305.




