Die Pflanzenschrift und ihre Offenbarungen
von Jagadis Chunder Bose
XXIII. KAPITEL
DIE NERVEN DER PFLANZEN
Wenn die Fingerspitze eines Menschen leicht gereizt wird, so entsteht dadurch ein Sinneseindruck, der im Gehirn als Empfindung wahrgenommen wird. Die Botschaft wird vermittelt längs eines Nervenstranges, welcher die Bahn für die Leitung der äußeren Sinneseindrücke bildet; wenn der Nerv auf irgendeine Weise beschädigt wird, so hört alle Empfindung auf. Die Reizung der Fingerspitze und die dadurch bewirkte Empfindung scheinen gleichzeitig zu sein, doch in Wirklichkeit bedarf es einer kurzen Zeit, um den Sinneseindruck von der Fingerspitze zum Gehirn zu leiten. Die Geschwindigkeit, mit der der Nerv den Eindruck vermittelt, kann etwa in folgender Weise festgestellt werden: die Versuchsperson gibt ein Zeichen, wenn sie einen bestimmten Reiz an der Zehe empfindet. Der Intervall, der zwischen der Reizung und dem Zeichen vergeht, befähigt uns, die Geschwindigkeit zu berechnen, mit der der Impuls die Länge des Nerven zurücklegt.
Wenn der Nerv an einem Muskel endigt, dann wird die Ankunft des Impulses durch die Zuckung des Muskels angezeigt. Experimente über Nervenimpulse werden gewöhnlich am Nerv und Muskel des Frosches ausgeführt, die im isolierten Zustand etliche Stunden lang am Leben erhalten werden können. Wenn man hier einen abliegenden Punkt des Nerven elektrisch reizt, so wird ein Impuls längs des Nerven bis an den Muskel geleitet, welcher an einem Registrierhebel befestigt ist. Die Registrierung erfolgt auf einer beweglichen Schreibtrommel, auf der auch durch einen Chronographen Zeitmarken verzeichnet werden. Die Geschwindigkeit der Reizleitung ergibt sich aus der Länge des Nerven und der auf der Trommel ablesbaren, für die Übertragung benötigten Zeit. In der Nervenbahn des Tieres lassen sich drei verschiedene Teile unterscheiden. Der erste ist der "Empfänger", der den äußeren Reiz aufnimmt; der zweite ist der "Leiter", der eigentliche "Nerv", der die Erregung auf gewisse Entfernung weitergibt, wobei indes keine sichtbare Veränderung in dem leitenden Gewebe während der Übertragung des Impulses zur Beobachtung kommt. Schließlich trifft der Impuls auf das reagierende Endorgan, den "Effektor" auf, der ein Muskel sein kann; die Reaktion wird da durch eine Bewegung sichtbar.
Die Anfänge eines solchen Nerven- und Muskelsystems sehen wir bei sehr niederen Tieren, wie bei der Seeanemone, wo eine Reizung der Tentakel an einem entfernten Körperteil eine Bewegungsreaktion bewirkt, ohne daß in der zwischenliegenden Partie eine Bewegung sichtbar ist. Der Empfänger und der Effektor liegen also in gewisser Entfernung, und der Nerv bildet das Verbindungsglied.
Diese Art der Erregungsübertragung, bei der die Wirkung des an einer Stelle eintreffenden Reizes sich durch eine Bewegung an einer anderen, abliegenden Stelle kundgibt, scheint derjenigen nicht unähnlich, die uns bei der Sinnespflanze Mimosa pudica entgegentritt. Hier veranlaßt ebenfall der auf einen Blättchenstiel einwirkende Reiz, der z. B. ein elektrischer sein kann, einen Impuls, welcher, längs des Blattstieles wandernd, das motile Organ, das Gelenkpolster erreicht, dessen Kontraktion das plötzliche Fallen des Blattes bewirkt. Wiewohl die Effekte bei der Pflanze und beim Tiar sehr ähnlich sind, so geht doch die herrschende Meinung dahin, daß der Reizimpuls sich bei der Pflanze in ganz anderer Weise weitergibt als im tierischen Nerv.
WASSERROHR ODER NERV?
Wir wollen uns umsehen, auf was für experimentelle Tatsachen dieser Schluß sich stützt. Pfeffer versetzte der Pflanze Messerschnitte, um sie zu reizen und zu beobachten, wie sie auf solche brutale Behandlung antwortet. Können wir uns vorstellen, daß ein Mensch eine normale Antwort gäbe, wenn er durch einen Messer hieb gereizt worden ist? Weit entfernt davon, zusammenhängend zu antworten, würde er nur vielleicht in Zuckungen verfallen! Nach Anbringung der Schnittwunde beobachtete Pfeffer an der Pflanze .den Austritt von Saft aus der Wunde. Den turge-szenten Stengel der Pflanze dachte er sich ähnlich einem mit Wasser gefüllten Kautschukschlauch; der Saftaustritt, so meinte er, sollte eine plötzliche Verminderung des Druckes bewirken und dadurch sollte ein Zug auf das sensitive Gelenkpolster ausgeübt werden. Dieser hydromechanische Impuls wird als praktisch gleichartig mit der Flüssigkeitsbewegung in einer Röhre betrachtet.
Ricca befreundet sich ebenfalls mit der Methode, Messerschnitte als Reizmittel anzuwenden. Er denkt sich, daß die Verwundung die Abscheidung irgendeiner stimulierenden Substanz, eines Hormones, bewirke und daß dieses hypothetische Hormon durch den aufsteigenden Saftstrom dem Blatte zugeführt werde und dessen Bewegung veranlasse. Es ist dies eine verfehlte Anwendung der Theorie der Hormone, wie sie von Starling und Bayliss ausgesprochen worden ist; denn diese Autoren haben ausdrücklich den grundlegenden Unterschied der zwei Arten der Kommunikation zwischen zwei entfernten Organen betont, die durch Stoffübertragung oder Bewegungsübertragung geschehen kann. Ein Beispiel der ersten Art bietet die langsame Bewegung von Flüssigkeiten, welche chemische Stimulantien in Lösung enthalten; wir begegnen solchen Fällen beim Saftaufstieg in der Pflanze oder bei der Blutzirkulation des Tieres. Die zweite Art ist die rasche Erregungsleitung von einem Punkte zum anderen, die mit der Fortpflanzung eines Nervenimpulses verknüpft ist. Man hat die beiden Arten der Übertragung nicht unpassend mit der Post und dem Telegraphen verglichen. Der Unterschied der Geschwindigkeit in beiden Fällen ist so groß, daß es ein unverzeihlicher Mißgriff wäre, die Fälle zu verwechseln.
Die hydromechanische Theorie und die Theorie vom Hormontransporte basieren beide auf der Meinung, daß eine Verwundung nötig ist, um eine mechanische Störung oder die Sekretion eines Reizstoffes zu bewirken. Beide sind zu verwerfen, wenn sich zeigen läßt, daß ein Erregungsimpuls in der Pflanze auch schon durch einen schwachen Reiz und ohne Verwundung erzeugt und weitergeleitet werden kann.
Die Pflanze ist in hohem Grade erregbar, und schon ein sehr schwacher Reiz reicht hin, einen Impuls auszulösen. Die einzige Entschuldigung dafür, einen Messerschnitt als Reiz anzuwenden, läge in der irrigen Annahme, daß die Pflanzen bei weitem weniger sensitiv wären wie die Tiere und daß sie deshalb mit Gewalt zur Tätigkeit angespornt werden müßten. Dies ist eine willkürliche und gänzlich unbegründete Annahme; habe ich doch bei Mimosa, wie früher ausgeführt, die Entdeckung gemacht, daß Erregung schon durch einen elektrischen Reiz bewirkt werden kann, der zehnmal schwächer ist als derjenige, den die menschlichen Sinne empfinden. Der Reiz bewirkt keine Verwundung, und doch wird die Erregung auf erhebliche Entfernung geleitet. Diese Tatsache allein beweist zur Genüge, wie gänzlich unbegründet sowohl die hydromechanische Theorie als auch die Theorie des Hormonaufstieges ist. Ich werde im folgenden noch weitere entscheidende Versuche beschreiben, die geeignet sind, diese Theorien gänzlich zu entkräften.
DIE THEORIE DER SAFTBEWEGUNG
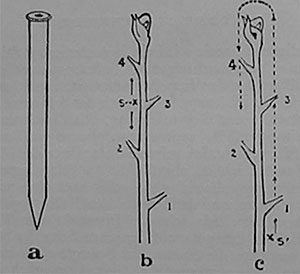 |
| Fig. 104. Die Wirkung einseitiger Reizung durch Ritzen, a) Der Ritzstift; b) Wirkung einer Reizung von mäßiger Stärke; der Reiz, der bei s, links, einwirkt, veranlaßt eine gleichzeitige Impulswelle nach oben und unten; c) Wirkung eines starken Reizes bei s', rechts; er verursacht einen Impuls, der rechts bis zur Spitze emporsteigt und sich dann links nach unten bewegt. |
Nach dieser Theorie wird ein hypothetisches Stimulans dem Blatte mit dem aufsteigenden Saftstrom zugeführt. In diesem Fall müßte der Impuls immer nach aufwärts fortschreiten, in der Richtung, in der sich der Saft bewegt, und nicht abwärts in der Gegenrichtung. Die Geschwindigkeit der Reizleitung müßte zudem etwa ebenso groß sein wie die der Saftbewegung. Ich reizte die eine Seite des Mimosastengels durch leichtes, oberflächliches Ritzen und fand, daß die Erregungswelle sich gleichzeitig nach oben wie nach unten fortpflanzte und beiderseits die Fallbewegung der Blätter veranlaßte. Als ich die Intensität der Reizung steigerte, lief die Erregung auf der einen Seite des Stengels aufwärts, erreichte die Spitze und bewegte sich dann auf der anderen Seite abwärts (Fig. 104). Unmöglich konnte der ansteigende Saftstrom solche charakteristischen Reaktionen bewirkt haben. Daß die Erregung am Stengel aufwärts und wieder abwärts geleitet wird, kann nur aus der Gegenwart eines spezifischen, leitenden Gewebes erklärt werden, das wir eben als Nervengewebe bezeichnen wollen.
Genaue Messungen, die ich ausgeführt habe, zeigen ferner, daß die Geschwindigkeit der Erregungsleitung eine um etliche hundert Male raschere ist, als die des Saftaufstieges. Daß die Saftbewegung nicht das mindeste mit der Erregungsleitung zu tun hat, zeigt dann auch das folgende Experiment in schlagender Weise: ein Tropfen Salzsäure wurde auf die Spitze des allerobersten Blattes einer Mimosa aufgetragen. Der erzeugte Impuls verbreitete sich auf erhebliche Entfernung nach abwärts, also entgegen der Richtung des normalen Saftstromes; die folgende chemische Untersuchung zeigte, daß die reizende Salzsäure nicht transportiert worden, sondern auf den Punkt der Einwirkung beschränkt geblieben war.
Nachdem ich so gezeigt habe, daß die Erregungsübertragung weder hydromechanischer Natur ist, noch auf der Saftbewegung beruht, will ich nun im folgenden klar zu machen suchen, daß die Reizleitung in der Pflanze auf der Fortpflanzung einer protoplasmatischen Erregungswelle beruht, - gerade so wie in dem erregten Nerv des Tieres.
BEWEISE FÜR DIE NERVÖSE NATUR DES IMPULSES
Es liegt auf der Hand, daß die mechanische Bewegung des Wassers durch eine Röhre innerhalb gewisser Grenzen nicht durch Hitze und Kälte beeinflußt werden wird; Anästketika werden die Röhre nicht bewußtlos machen und die Wasserströmung hemmen, und die Leitfähigkeit der
Röhre wird nicht vernichtet werden, wenn man sie mit einer Binde umgibt, die mit einem Gift getränkt ist. Diese Mittel werden hingegen die Übertragung einer nervösen Erregung tiefgreifend beeinflussen. Ob ein Impuls mechanischer oder nervöser Natur ist, läßt sich auf verschiedene Weise mit Sicherheit entscheiden.
Wenn Veränderungen der physiologischen Außenbedingungen die Geschwindigkeit der Reizleitung beeinflussen, dann muß der Impuls nervösen Charakter haben; fehlen dagegen solche Wirkungen, so wird der mechanische Charakter des Impulses erwiesen.
Es gibt verschiedene physiologische Mittel, die es gestatten, den nervösen Impuls zu verlangsamen oder aufzuhalten, ohne auf den mechanischen Impuls gleichartige Wirkungen auszuüben. Als Beispiele seien die folgenden genannt:
1. Wenn das leitende Gewebe oder der Nerv abgekühlt wird, so wird die Geschwindigkeit des Impulses herabgemindert und die Leitung kann endlich ganz still stehen.
2. Giftige Lösungen, die auf den Nerven einwirken, vernichten sein Leitvermögen dauernd.
3. Das Leitvermögen wird vorübergehend aufgehoben, wenn ein elektrischer Strom einen Teil der Nervenbahn durchfließt, durch die der Impuls sonst geleitet würde. Dieses "elektrotonische" Hindernis wird beseitigt, wenn der Strom zu fließen aufhört.
MESSUNG DER GESCHWINDIGKEIT DER REIZLEITUNG
 |
| Fig. 105. Bestimmung der Geschwindigkeit der Erregungsleitung im Blattstiel von Mimosa. Unten zwei Reaktionskurven auf indirekte Reizung im Abstand von 30 mm; oben die Reaktionskurve auf direkte Reizung, sie veranschaulicht die Latenzzeit. 10 Vibrationen in der Sekunde. |
Die Versuche wurden durch automatische Bestimmung der Geschwindigkeit ausgeführt, mit der der Impuls erstens unter normalen Bedingungen und sodann unter veränderten Bedingungen solcher Art fortschreitet, wie sie die Erregungsleitung im tierischen Nerv beeinflussen. Die normale Geschwindigkeit ergibt sich, indem man einen elektrischen Reiz von bestimmter Stärke auf den Blattstiel von Mimosa in bestimmtem Abstand vom Gelenkpolster, von z. B. 30 mm einwirken läßt, wobei die Reizintensität in einer Folge von Versuchen konstant erhalten werden kann. Der Moment der Reizeinwirkung ist in der Figur 105 durch eine vertikale Linie verzeichnet. Der "Resonant-Recorder" zeichnet seine Marken in bestimmten Intervallen von z. B. je einer Zehntel-Sekunde auf und mißt so die Zeit, die zwischen der Einwirkung des Reizes und der Reaktionsbewegung des Blattes, welcher der Anstieg der Kurve entspricht, vergeht. Der Intervall zwischen der Reizung und der Reaktion ergibt sich zu 16,2 Einheiten, deren jede 0,1 Sekunden ausmacht (Fig. 105). Der ganze Intervall beträgt somit 1,62 Sekunden. Um die Zuverlässigkeit der Methode zu prüfen, wurden nacheinander zwei Registrierkurven aufgenommen; sie zeigen, daß die unter normalen Bedingungen verstreichende Zeit beide Male praktisch die gleiche ist. Die gemessene Zeit schließt auch die Latenzperiode des Gelenkpolsters ein, entsprechend der Zeit, die vergeht, um den motorischen Mechanismus in Bewegung zu setzen. Die Dauer der Latenzperiode, d. h. der Zeit von der Reizung bis zur Reaktion, ergibt sich, wenn der Reiz direkt auf das Gelenkpolster einwirkt. Sie war im vorliegenden Fall 0,12 Sekunden und die tatsächliche Dauer der Reizleitung auf eine Entfernung von 30 mm ist daher 1,62-0,12 Sekunden oder 1,5 Sekunden. Die Geschwindigkeit der Reizleitung ist daher 30/1,5=20 mm pro Sekunde.
Die Geschwindigkeit der Erregungsübertragung ändert sich mit dem physiologischen Zustand der Pflanze. Sie ist im Sommer größer als im Winter. Eine auffällige Beobachtung ist, daß dicke Exemplare nur gemächlich reagieren, während bei dünnen Exemplaren der Gipfel der Erregung in unglaublich kurzer Zeit erreicht wird. Derartige Unterschiede sind selbst beim Menschen nicht unbekannt. In einem dünnen Blattstiel von Mimosa kann die Geschwindigkeit 400 mm pro Sekunde oder 24000 mm pro Minute erreichen. Die Geschwindigkeit der Reizleitung bei Mimosa ist zwar kleiner als jene bei den höheren Tieren, doch beträchtlich größer als jene bei den niederen Tieren, z. B. bei Anodon. Die Geschwindigkeit bei der Pflanze kann somit als ein Mittelglied zwischen jenen betrachtet werden. Die Geschwindigkeit der Reizleitung nimmt bei Mimosa und beim tierischen Nerv in gewissen Grenzen mit dem Steigen der Temperatur zu, mit dem Fallen ab. Ein Temperaturanstieg von etwa 9° verdoppelt bei Mimosa die Geschwindigkeit.
LEITUNGSHEMMUNG DURCH EIN PHYSIOLOGISCHES HINDERNIS
Man kann ein Teilstück der Bahn, die der Impuls zu durchwandern hat, in solcher Weise behandeln, daß die Leitung verzögert oder ganz aufgehalten wird (Fig. 106). Hemmung durch Kälte. - Wird ein Stück des Blattstieles von Mimosa durch kaltes Wasser mäßig abgekühlt, so zeigen die Registrierkurven, daß die Reizleitungszeiten verlängert werden. Extreme Abkühlung durch eiskaltes Wasser vernichtet die Leitfähigkeit gänzlich (Fig. 107). Um zu zeigen, daß die lokale Kälteeinwirkung nur das Leitvermögen vernichtet hatte, ohne die Erregbarkeit des Gelenkpolsters zu beeinflussen, wurde der letztere direkt gereizt; er wies normales Reaktionsvermögen auf.
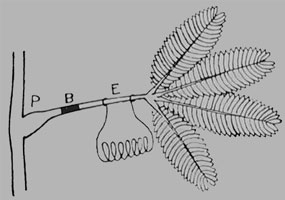 |
 |
| Fig. 106. Die verschiedenen physiologischen Hindernisse (Blocks) wirken bei B auf die Leitungsbahn ein. |
Fig. 107. Kälte bewirkt Verlangsamung und Stillstand der Reizleitung. (1) Normale Kurve; (2) Verlangsamung bei gelinder Abkühlung; (3) Aufhören der Reizleitung infolge intensiver Kälte; (4) die Kurve bei direkter Reizung. |
Mit diesen Versuchen beschäftigt, lernte ich die interessante Erscheinung kennen, daß als Nachwirkung intensiver Kälte die Reizleitung gelähmt wird und daß diese Lähmung länger als eine Stunde andauert, auch nachdem das Gewebe wieder auf normale Temperatur gebracht worden ist. Elektrische Reize werden wirksam darin befunden, gewisse Formen der Lähmung beim Menschen zu heilen, und ich entdeckte die recht auffällige Tatsache, daß das verlorene Reizleitungsvermögen bei der Pflanze rasch wieder hergestellt werden kann, wenn man den gelähmten Teil des Blattstieles der Wirkung tetanisierender elektrischer Reize unterzieht.
Hemmung durch Gift. - Ich will weiter zeigen, wie Gifte eine dauernde Vernichtung des Leitungsvermögens bewirken. Ein 1,5 cm breiter Stoffstreifen wurde um einen leitenden Blattstiel gewunden, und eine giftige Lösung von Kaliumcyanid aufgetragen. Die Wirkung war so stark, daß das Leitvermögen bereits innerhalb einer Zeit von fünf Minuten vernichtet war. Obwohl die Intensität des Reizes etwa auf das Achtfache gesteigert wurde, erhielt ich keine Reaktion. Direkte Reizung des Gelenkpolsters zeigte, daß dessen Bewegungsvermögen keine Veränderung erlitten hatte (Fig. 108).
 |
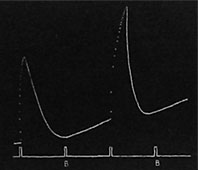 |
| Fig. 108. Vernichtung der Leitfähigkeit durch Kaliumcyanid. (1) Normale Kurve; (2) Aufhören der Reizleitung nach 5 Minuten langer Einwirkung; (3) Kurve, die das Fehlen des Impulses selbst nach starker Reizung zeigt; (4) die Kurve bei direkter Reizung. |
Fig. 109. Aufzeichnung der Erregungsleitung mit und ohne Hindernis. Stillstand der Erregungsleitung durch den elektrotonischen Block bei B, B. |
Elektrotonische Hemmung. ("Elektrotonischer Block.") - Im obigen Fall wurde die Leitfähigkeit dauernd vernichtet; beim tierischen Nerv wird, wie erwähnt, das Leitvermögen vorübergehend aufgehoben, wenn man einen konstanten elektrischen Strom durch einen Teil der Leitbahn schickt. Das Hindernis besteht solange, als der elektrische Strom fließt, und das Leitvermögen wird sogleich wieder hergestellt, wenn der Strom unterbrochen wird. Ganz analoge Resultate erhielt ich bei Mimosa; der hemmende Strom wurde (Fig. 109) bei B geschlossen und nachher wieder unterbrochen. Dies wurde zweimal wiederholt und die Kurve zeigt, daß die Erregungsleitung jedesmal unterdrückt war, solange der hemmende Strom einwirkte, und daß sie nach dem Aufhören des Stromes wieder hergestellt wurde.
POLARE ERREGUNG DURCH ELEKTRISCHEN STROM
Ich will nun noch Versuchsergebnisse mitteilen, welche den gleichartigen Charakter der Erregungsübertragung beim Tier und bei der Pflanze aufs klarste erkennen lassen.
 |
| Fig. 110. Das Bild links zeigt die Erregung beim Stromschluß an der Kathode, das Bild rechts jene bei der Stromöffnung an der Anode (Biophytum sensitivum). |
Es ist wohl bekannt, daß beim tierischen Nerv ein elektrischer Strom spezifische Erregungswirkungen auslöst. Wenn ein Strom plötzlich durch einen Nerv gesandt wird, dann tritt Erregung an dem Punkte des Nerven ein, wo die Kathode liegt, d. h. wo der Strom aus dem Gewebe austritt. Wenn der Strom plötzlich unterbrochen wird, so springt die Erregung nach der Anode über, d. h. der Stelle, wo der Strom in das Gewebe eintritt. Die Erregung bleibt nicht lokalisiert, sondern wird auf gewisse Entfernung geleitet, wie die Kontraktion eines Muskels am Ende des Nerven zeigt. Ein ganz ähnliches Ergebnis erhielt ich bei Mimosa und anderen sensitiven Pflanzen. Ich gebe eine Skizze, die jene Effekte bei Biophytum sensitivum zeigt (Fig. 110). Die Figur zur Linken läßt die Erregung erkennen, die der eintretende Strom am Kathodenpunkt auslöst; die Erregung breitete sich nach beiden Richtungen aus. Nachdem die Blättchen zur Ruhelage zurückgekehrt waren, wurde der Stromkreis unterbrochen, und die Erregung erschien nun am Punkte der Anode.
Indem also die Erregung beim Tier wie bei der Pflanze beim Stromschluß an der Kathode, bei der Öffnung an der Anode erscheint, da ferner der Impuls in beiden Fällen durch starke Kälte, durch Giftwirkung, durch elektrotoni-sche Wegsperre gehemmt wird, so ergibt sich unausweichlich der Schluß, daß die Übertragung in beiden Fällen im Wesentlichen auf dem gleichen physiologischen Prozeß beruht. Wird dieser beim Tier als "nervös" bezeichnet, so dürfen wir ihn mit gleichem Recht auch bei der Pflanze so benennen.




