Die Pflanzenschrift und ihre Offenbarungen
von Jagadis Chunder Bose
XVIII. KAPITEL
DAS BEI DER SAFTBEFÖRDERUNG TÄTIGE GEWEBE
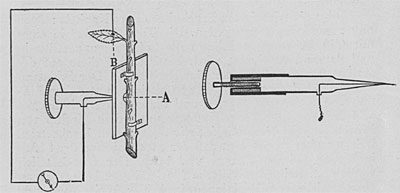 |
| Fig. 74. Die elektrische Sonde zur Bestimmung der Lage der pulsierenden Zellschicht. Die Spitze der Sonde dringt bei A in den Stengel ein, wobei der zweite elektrische Kontakt an einem entfernten Blatt hergestellt wird. Die Figur rechts stellt vergrößert die Sonde mit der Mikrometerschraube dar, die zur schrittweisen Einführung der Sonde in die Gewebe der Pflanze dient. |
Die im letzten Kapitel beschriebenen Versuche beweisen, daß irgendwo im Inneren der Pflanze ein aktiv tätiges Gewebe vorhanden ist, dessen Pulsationen die Fortbewegung des Saftes bewirken, gerade so wie die Pulsation des Herzens den Blutumlauf bei den Tieren unterhält. Es muß daher eine Art primitives "Herz" in der Pflanze sein, doch nicht so zentralisiert und hoch differenziert wie in höheren Tieren. Bei den niederer organisierten Tieren, und ebenso beim Embryo der höheren, ist das Herz eine verlängerte Röhre und die in ihm enthaltene Nährflüssigkeit wird durch peristaltische Kontraktionen vorwärts bewegt. Die Fortbewegung des Saftes bei den Pflanzen beruht nun nach meinen Befunden auf einer ähnlichen peristaltischen Wirkung. Da der pulsierende Mechanismus bei den Pflanzen nicht so hoch differenziert ist wie beim höheren Tier, so ist das Bewegungsorgan nur einem verlängerten "Herzen" vergleichbar, wenn man diesen Ausdruck im weiteren Sinne gebraucht. Man könnte sagen, daß säftebewegende System der Pflanze entspricht dem Herzen und den Arterien der Tiere.
DIE SUCHE NACH DEM "HERZEN" DER PFLANZE
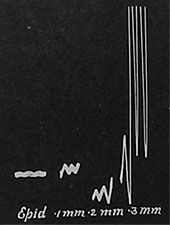 |
| Fig. 75. Registrierkurve der Amplitude der elektrischen Pulsationen in verschiedenen Gewebsschichten. Man beachte die plötzliche Zunahme in einer Tiefe von 0,3 mm unter der Oberfläche, die betreffende Zellschicht lag innen in der primären Rinde; ein Teil der Kurve liegt schon außerhalb der Platte. |
Wo liegt nun dieses "Herz"? Ist es möglich, die Lage der aktiven Zellen im Innern des Baumes genau zu bestimmen und sie bei ihrer wirklichen Pumptätigkeit zu erfassen? Bei derartigen Versuchen müssen wir bestrebt sein, einen Einblick zu gewinnen in die kleinste Lebenseinheit, in die individuelle Zelle oder das "Lebens-atom", und ihren Pulsschlag aufzuzeichnen. Die Pulsa-tionsbewegung der Einzelzelle ist ultramikroskopisch, und es mag scheinen, daß ihre Beobachtung jenseits der Grenzen des Möglichen liegt. Trotzdem ist die Lösung dieser Aufgabe möglich geworden und zwar durch meine "elektrische Probe", die ich früher bei meinen Studien über die Lage der Perzeptionsorgane für den Reiz der Schwerkraft angewandt hatte.
Die Lage des Herzens zu bestimmen, ist mir durch Anwendung der elektrischen Sonde gelungen, die mit dem Stromkreis eines empfindlichen Galvanometers verbunden war. Wird ein elektrischer Kontakt mit einem ruhenden Muskel hergestellt, so bleibt das Galvanometer in Ruhe. Wird der Kontakt aber mit einem schlagenden Herzen hergestellt, so erhält man elektrische Pulsationen, die den mechanischen Pulsationen entsprechen. Auf der Suche nach dem Herzen der Pflanze führte ich die Sonde langsam, Schritt für Schritt, in das Pflanzenorgan ein; sobald sie in Kontakt mit der pulsierenden Zellage kam, wurden elektrische Signale ausgesandt, die vom Galvanographen automatisch verzeichnet wurden.
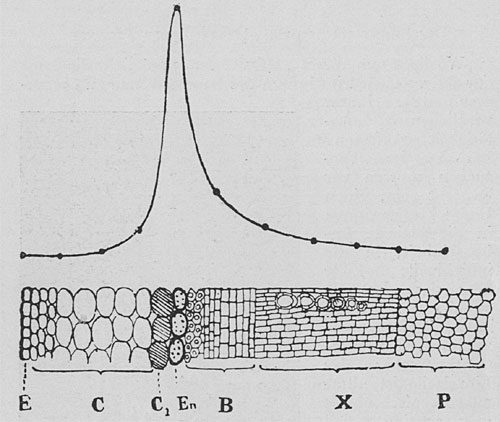 |
| Fig. 76. Schnitt durch den Blattstiel von Brassica und Kurve der Pulsationstätigkeit in verschiedenen Gewebsschichten. E Epidermis, C Rinde, Cj die aktive innerste Rindenschicht, En Endodermis, B Phloem, X Xylem, P Mark. Man beachte den plötzlichen Anstieg der Pulsationstätigkeit in der Zellschicht Q. |
 |
| Fig. 77. Aufzeichnung der Pulsationen beim Mangobaum. |
Der Versuch wurde in folgender Weise durchgeführt. Der eine Pol des Galvanometers wird mit einem ruhenden Gewebe, z. B. mit der Blattepidermis verbunden; der andere Pol wird mit der Sonde verbunden, die nun in der Querrichtung langsam in Schritten von 0,1 mm in den Stengel eingeführt wird. In der Epidermis ließ sich keine Pulsation nachweisen; als die Sonde eine Tiefe von 0,1 mm erreichte, zeigte sich eine schwache Pulsation; eine ähnliche fand sich in einer Tiefe von 0,2 mm. Als aber die Nadel 0,3 mm erreichte, da zeigte die Pulsation einen plötzlichen Anstieg; dieser war so groß, daß ein Teil der Kurve nicht mehr auf die Platte kam (Fig. 75); offenbar war die Nadel in Berührung mit den pulsierenden Zellen gekommen. Als sie noch tiefer in den Stengel eingeführt wurde, da verschwand die Pulsationstätigkeit plötzlich wieder. Wenn nun ein Querschnitt durch den Stengel angefertigt wurde, der die Einstichlinie der Nadel enthielt, so ergab sich, daß die maximale Tätigkeit in dem Moment beobachtet worden war, wo die Nadel die innerste Schicht der primären Rinde, die an das junge Stranggewebe grenzt, getroffen hatte. Der Kontakt der Nadel mit dem Holz verursachte keine Pulsation, woraus hervorgeht, daß das tote Holz an der Fortbewegung des Saftes keinen aktiven Anteil nimmt. Eine Darstellung der verschiedenen Gewebslagen im Blattstiele von Brassica mit ihrer ungleichen Pulsationstätigkeit gibt die Fig. 76. Die Kurve zeigt, daß die Tätigkeit bei C1 in der Innenrinde am stärksten ist. Die Lichtmarke, die vom Galvanometer reflektiert wird, läßt durch ihre Schwingungen nach rechts und links die unsichtbaren Pulsationen der aktiven Zellen im Inneren der Pflanze sichtbar werden. Fig. 77 zeigt die Registrierkurve der elektrischen Pulsationen der aktiven Zellschicht des Mangobaumes.
BEWEISE FÜR DIE HERZTÄTIGKEIT
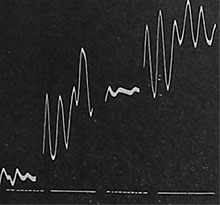 |
| Fig. 78. Aufzeichnung der elektrischen Pulsationen bei abwechselnder Trockenheit und Wasserzüfuhr. Die unterbrochene Linie unten bedeutet Trockenheit, die zusammenhängende Linie erneute Wasserzufuhr. |
Was für Beweise haben wir dafür, daß diese elektrischen Pulsationen wirklich eine Pulstätigkeit, ähnlich der des Herzens, in der Pflanze anzeigen? Es gibt eine Reihe von Merkmalen, welche die spezifische Tätigkeit des Herzens kennzeichnen; es wird genügen, die folgenden aufzuzählen: 1. Der Herzschlag kommt zum Stillstand, wenn der innere Blutdruck gering ist, die Pulsationen beginnen wieder, wenn der Innendruck zunimmt. 2. Desgleichen steht der Pulsschlag still, wenn das Herz sich in einem Depressions- oder subtonischen Zustand befindet; eine Stimulation kann den Herzschlag dann neu beleben. 3. Anästhetika, wie Chloroform, stimulieren im Beginn den Herzschlag, doch bei andauernder Anästhesierung steht dieser still, worauf der Tod des Tieres folgt. Ich will nun zeigen, daß die Reaktionen in der Pulsationstätigkeit bei der Pflanze denjenigen beim Tier durchaus parallel verlaufen.
DIE WIRKUNG VERMINDERTEN UND ERHÖHTEN INNENDRUCKES
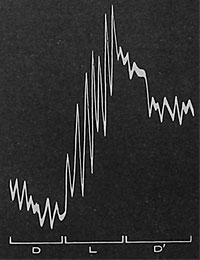 |
| Fig. 79. Die Wirkung des Lichtreizes; sie besteht in der Beschleunigung der elektrischen Zellpulsation eines subtonischen Objekts. D schwache Pulsation im Dunkeln; L verstärkte Pulsation bei Belichtung, D'Depression bei erneuter Verdunkelung. |
Der Saftdruck erfährt bei Trockenheit eine starke Verminderung, wodurch die Pulsationen der bei der Saftbewegung tätigen Zellschicht zum Stillstand kommen. Wenn der Saftdruck nach dem Begießen wieder zunimmt, so lebt die stillgelegte Pulsationstätigkeit wieder auf. Ich gebe eine Kurve wieder (Fig. 78), welche die Wirkung abwechselnden Wasserentzuges und Begießens auf die elektrischen Pulsationen der aktiven Zellschicht zeigt. Man sieht, wie die Pulsationen mehrmals nacheinander durch Trockenheit herabgedrückt werden und nach der Zufuhr von Wasser wieder aufleben.
DIE WIRKUNG SUBTONISCHEN ZUSTANDES
Wenn eine Pflanze durch 24 Stunden dunkel gehalten wird, so wird ihr Zustand so stark subtonisch, daß sie die Fähigkeit, den Saftaufstieg zu unterhalten, verliert. Eine Stimulation durch Licht oder durch elektrische Reizung kann dann die Saftsteigetätigkeit neu beleben. Was mag in diesem Fall die wahre Ursache der Veränderung sein? Die Erklärung ergibt sich aus den Registrierkurven der elektrischen Pulsationen. Diese zeigen, daß, während die Pulsationen unter der Wirkung dauernder Dunkelheit praktisch stille stehen, die Pulstätigkeit nach elektrischer Reizung oder Belichtung neu auflebt und nach dem Aufhören des Reizes neuerlich zum Stillstand kommt (Fig. 79).
WIRKUNG DER ANÄSTHETIKA
Es wurde gezeigt, wie Chloroform im ersten Stadium der Einwirkung die Tätigkeit des Herzens erhöht, während dauernde Einwirkung zur Depression und zum Stillstand führt. Ich habe nun gefunden, daß in gleicher Weise auch die Geschwindigkeit des Saftaufstieges unter dem Einfluß von Chloroform zunächst eine Beschleunigung zeigt, worauf dann eine Depression und ein völliger Stillstand folgt.
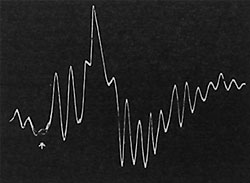 |
| Fig. 80. Wirkung des Chloroforms auf die Pulsationen, der Pfeil bezeichnet den Beginn der Einwirkung. Man beachte die anfängliche Verstärkung der Pulsationen, wobei die aufsteigenden Kurvenäste verlängert sind; nach fortgesetzter Einwirkung hörten die Pulsationen ganz auf. |
Die elektrische Pulsationskurve der bei der Saftbewegung tätigen Zellschicht zeigt unter der Wirkung von Chloroform gleichsinnige Reaktionen. Der anfänglichen Beschleunigung folgt eine Depression und ein Stillstand der Pulsations-tätigkeit (Fig. 80).
Diese Versuchsresultate liefern den Beweis dafür, daß das Saftsteigen durch die Pulsationstätigkeit der Zellen einer bestimmten Zellschicht bewirkt wird, und zwar der inneren Schicht der Rinde, die den Gefäßzylinder umschließt. Man darf in diesem Zusammenhange nicht vergessen, daß unter besonderen Umständen alle lebenden Zellen in einen Zustand rhythmischer Tätigkeit versetzt werden können. Gewisse Zellschichten besitzen aber naturgemäß eine stärkere Aktivität als andere und es beruht auf der Pulsationstätigkeit jener Zellen der inneren Rinde, daß der normale Saftaufstieg im Gang erhalten wird.
Die das Wasser befördernde Zellschicht ist im Dicoty-ledonenstamm ein Hohlzylinder, welcher den Stamm seiner ganzen Länge nach durchzieht. Dieser Zylinder grenzt unmittelbar an das junge Stranggewebe. Die Funktion dieses langgestreckten Zylinders ist, wie bereits erklärt, nicht unähnlich der des verlängerten Herzens gewisser niederer Tiere, in welchem die Blutzirkulation durch eine Folge peristaltischer Wellen hervorgebracht wird. Die Saftbeförderung bei den Pflanzen hat sich als ein im Wesen ähnlicher peristaltischer Prozeß erwiesen, wobei jede Kontraktionswelle den Saft nach vorwärts treibt. Eine Folge solcher peristaltischer Wellen hält den Saftaufstieg beständig im Gang.
NORMALE UND UMGEKEHRTE PERISTALTISCHE WELLEN
Man ist vielleicht geneigt zu fragen, warum denn die peristaltischen Wellen den Saft gerade immer aufwärts treiben. Wäre es wohl möglich, die Richtung jener Wellen umzukehren, so daß sie den Saft nach abwärts fließen lassen? Was für Bedingungen bestimmen die Richtung des Saftflusses?
Das allgemeine Gesetz für die Richtung, in der der Saft sich bewegt, möchte ich so formulieren, daß er stets von der aktiveren zur minder aktiven Region hin fließt. Es ist klar, daß die Pulsationstätigkeit, wenn sie an den beiden Enden eines Organes gleich stark wäre, sich die Wage halten würde, und daß dann keine Bewegung des Saftes nach der einen Richtung resultieren könnte. Ein Unterschied in der Intensität der Bewegung an den zwei Enden des Organes kann auf zweierlei Weise zustande kommen: erstens durch Turgordifferenzen und zweitens durch differente Reizung. Ich will erklären, was unter Turgordifferenz und differenter Reizung zu verstehen ist.
Nach einer Zufuhr von Wasser wird das untere Ende eines Baumes durch Wasseraufnahme prall und turgeszent, wogegen am oberen Ende infolge des starken Wasserentzuges durch die transpirierenden Blätter das Welken beginnt. Nun haben wir gesehen, daß die rhythmische Tätigkeit der das Wasser befördernden Zellen durch Turgor-zunahme erhöht, durch Trockenheit geschwächt wird. Die Saftbeförderung erfolgt daher von der stärker turgeszenten, aktiveren zu der minder turgeszenten, inaktiven Region. Die Saftbewegung folgt somit dem "Turgorgefälle", sie strebt, den Grad der Turgordehnung in den verschiedenen Teilen der Pflanze auszugleichen.
Ich will nun zeigen, daß es möglich ist, die normale Aufwärtsrichtung des Saftstromes in die umgekehrte zu verwandeln, indem man einfach das Turgorgefälle umkehrt. Wird beispielsweise einer Topfpflanze das Wasser entzogen, so wird der Stengel bald durch das Austrocknen schlaff, er hängt über und die welkenden Blätter hängen herab. Die Pulsationstätigkeit ist jetzt in der ganzen Pflanze unterbrochen. Wird nun ein Glas Wasser derart aufgestellt, daß der oberste Teil des welken Stengels eintaucht, dann nimmt der obere Teil des Stengels Wasser auf und wird dadurch stärker turgeszent als das untere Ende. Die Pulsationstätigkeit wird am oberen Ende neu belebt; das Turgorgefälle wird umgekehrt, und der Saft fließt nun abwärts, entgegen der Richtung der normalen Strömung. Dieser umgekehrte Saftstrom wird dadurch ersichtlich, daß die welken Blätter vom Gipfel des Stengels nach abwärts nacheinander wieder frisch werden. Ich habe die Geschwindigkeit des normalen, aufsteigenden Saftstromes und die des umgekehrten gemessen; das Ergebnis war, daß die Geschwindigkeit in der umgekehrten, ungewohnten Richtung viel kleiner ist. Bei der Blutzirkulation wird die Vorwärtsbewegung durch das Vorhandensein von Klappen unterstützt, welche die Strömung nach der einen Richtung hin begünstigen. Die Scheidewände der Zellpumpen in der Pflanze wirken wohl irgendwie ähnlich dahin, die Saftbewegung in der normalen Aufwärtsrichtung zu begünstigen. Unter normalen Umständen erfahren die Wurzelzellen durch die Reibung an den Bodenteilchen eine beständige Reizung, und dies gibt vermutlich den Anlaß für die Entstehung der peristaltischen Wellen. Auch die Turgorsteige-rung am unteren Ende bei Wasserzufuhr verursacht eine Saftbeförderung von unten nach oben, von der turgeszen-teren zur minder aktiven Region.
FUNKTION DES HOLZES
Bei den krautigen Pflanzen ist der Abstand der Blätter vom Bodenwasser nicht allzu groß, doch bei den hohen Bäumen ist es nötig, daß eine den Blättern nähere Wasserquelle, eine "Erweiterung des Bodenwassers" vorhanden ist, und diese besteht in wassergefüllten Röhren. Diese Wasserleitungsröhren sind die jungen Holzgefäße im Splint, welche der mechanischen Beförderung des Wassers in der Zeit der lebhaften Transpiration der Blätter dienen. Wenn die Transpiration schwach ist, so versorgt der normale Saftaufstieg längs der Rinde alle Teile des Baumes mit Wasser; die Blätter werden turgeszent, und das Splintholz füllt sich mit Wasser. Während lebhafter Transpiration aber reicht die physiologische Saftbeförderung nicht aus, den Bedarf zu decken, und es wird aus den Holzgefäßen Wasser entnommen. Dann treten also zwei Faktoren in Tätigkeit - die physiologische Saftbeförderung vermittels der aktiven Rindenzellen und längs dieser und der physikalische Transport längs der Holzgefäße im Xylem.
Wir wollen nun die wichtigsten Glieder des Saftsteigeprozesses nochmals ins Auge fassen. Die absorbierenden Wurzelzellen werden beständig durch mechanische Reibung an den Bodenteilchen gereizt, und dies gibt Anlaß zur Entstehung peristaltischer Pulsationswellen längs der aktiven, saftbefördernden Zellschicht der inneren Rinde. Die Richtung des Saftstromes wird durch das Turgorgefälle bestimmt, durch die Differenz zwischen dem stärker turgeszenten, unteren Teil der Pflanze und dem oberen Teil, der während der lebhaften Transpiration der Blätter dem Welken nahe ist. Die rhythmischen Kontraktionsbewegungen der aktiven Zellen treiben den Saft nicht nur aufwärts, sondern sie pumpen ihn auch in seitlicher Richtung in das junge Xylem, welches als Reservoir für die Zeiten erhöhten Bedarfes funktioniert; ihm wird Wasser entnommen, während die Transpiration am lebhaftesten ist.




