Die Pflanzenschrift und ihre Offenbarungen
von Jagadis Chunder Bose
XVII. KAPITEL
DAS SAFTSTEIGEN
Die Pflanze nimmt Wasser aus dem Boden auf, in welchem Nährstoffe gelöst sind. Die Verteilung der absorbierten Flüssigkeit, das Saftsteigen, befähigt die Pflanze, die Zellen im turgeszenten Zustand zu erhalten, ohne welchen das Wachstum der Pflanze und ihre aktiven Bewegungen nicht möglich wären. Wie wird aber der Saft bis zur Spitze des Baumes emporgehoben?
Diese Frage hat die Forscher seit mehr als zweihundert Jahren in Atem gehalten. Man hat einerseits angenommen, daß das Saftsteigen allein auf der Wirkung physikalischer Kräfte beruht, andererseits, daß es durch eine aktive Tätigkeit der lebenden Gewebe hervorgebracht wird. Aus Gründen, die gleich dargelegt werden sollen, vereinigt sich derzeit die Mehrzahl der Stimmen zugunsten der physikalischen Theorie, doch die im folgenden mitzuteilenden Erfahrungen zeigen, daß diese Auffassung ganz unhaltbar ist. Betrachten wir z. B. einen großen Baum mit zahlreichen Blättern. Was für physikalische Kräfte können wir hier zur Erklärung der Hebung des Saftes von der Wurzel bis zu den Blättern heranziehen? An den Blättern findet fortwährend Transpiration, d.h. Verdunstung von Wasserdampfstatt und dadurch entsteht in den Holzgefäßen, welche die Pflanze vom Grunde bis zum Gipfel durchziehen, ein teilweises Vakuum. Der äußere Luftdruck wird infolgedessen dahin wirken, das Wasser in der Pflanze zu heben; doch die größte Höhe, bis zu der Wasser gehoben werden kann, beträgt 10 m, d. i. die Höhe des Wasserbarometers. Die Palmen wachsen indes oft bis zu einer Höhe von 30 m; doch die Palme ist noch ein Zwerg im Vergleich zu gewissen Riesen des Pflanzenreiches, wie Eucalyptus amygdalina mit einer Höhe von 150 m. Die Theorie vom Luftdruck ist daher unzureichend und gleiches gilt von der Kapillaritätstheorie.
Auch die osmotische Theorie ist zur Erklärung der Erscheinungen herangezogen worden. Wenn eine semipermeable Zelle, die eine starke Zuckerlösung enthält, in Wasser gestellt wird, so tritt Wasser in die Zelle mit der Zuckerlösung ein. Die Pflanzenzellen verhalten sich ähnlich wie gewisse künstliche Zellen, die mit starken Lösungen erfüllt sind, und sie "werden daher Wasser, das ihnen zugänglich ist, aufnehmen. Die Zellen der Wurzel nehmen Wasser aus dem Boden auf, die nächst höheren Zellen entnehmen es den tiefer gelegenen und dieser Vorgang setzt sich nach oben hin weiter fort.
Diese osmotischen Vorgänge verlaufen nun aber äußerst langsam. Laßt uns sehen, was dies bedeutet, wenn etwa die Blätter am Gipfel eines Eukalyptusbaumes infolge starker Trockenheit dem Tode nahe sind. Wir können uns wohl vorstellen, mit welcher Ungeduld das vertrocknende Laub den aufsteigenden Saft erwarten mag, wenn endlich ein Regen die Wurzeln befeuchtet hat. Würde die Wasserbewegung allein auf osmotischer Wirkung beruhen, dann wäre die Aussicht für die Blätter keine allzu erfreuliche, denn die kürzeste Zeit, in der das Wasser durch Osmose bis zum Gipfel gelangte, würde wesentlich mehr als ein Jahr betragen! Unter den Verteidigern der physikalischen Theorie steht Strasburger, ein hervorragender Pflanzenphysiologe; doch selbst er war gezwungen zu sagen: "Die osmotischen Strömungen verlaufen zu langsam, um hier in Betracht zu kommen, und es fehlt außerdem die für eine solche Bewegung notwendige, bestimmte Verteilung osmotisch wirksamer Stoffe."4 Es müssen daher noch irgendwelche andere Kräfte am Werk sein, eine raschere Bewegung des Saftes herbeizuführen.
ZUG- UND DRUCKWIRKUNG DER ENDORGANE
Eine andere Theorie sagt aus, daß das Saftsteigen auf der Wirkung eines Zuges von oben und eines Druckes von unten beruht, den die Organe an den betreffenden Enden, die Blätter und die Wurzel, ausüben. Diese Theorie ist nur zum Teil eine physikalische, denn sowohl der Zug als auch der Druck kommen durch die aktive Tätigkeit der lebenden Blätter, resp. der Wurzel zustande. Man nimmt an, daß die Wasserentziehung, welche die Blätter durch die Transpiration erfahren, einen Zug längs der kohärenten Wassersäulen in den Holzgefäßen ausübt. Aber die Wassersäulen in den Gefäßen sind nicht zusammenhängend, sondern sie sind durch Luftblasen unterbrochen; es ist offenbar unmöglich, Wasserschnüre zur Erklärung des Transportes heranzuziehen, z.umal wenn diese noch stellenweise unterbrochen sind! Soviel zum Zug von oben. Der Druck von unten wird, so nimmt man an, durch den Wurzeldruck ausgeübt. Doch bei der Palme besteht kein nachweisbarer Wurzeldruck, und doch steigt der Saft bis zu einer Höhe von mehr als 30 m. Ferner ist zur Zeit der lebhaften Transpiration, wenn das Wasserbedürfnis des Baumes am größten ist, der Wurzeldruck nicht positiv, sondern negativ. Wir werden auch später noch nachweisen, daß weder das Blatt noch die Wurzel für die Weiterbewegung des Saftes absolut notwendig ist, indem ich zeigen werde, daß die Saftbewegung selbst nach völliger Entfernung der Wurzel und der Blätter fortdauert.
DIE SAFTBEWEGUNG BERUHT WESENTLICH AUF VITALER TÄTIGKEIT
Da sich die verschiedenen physikalischen Theorien als unzureichend erwiesen haben, so verbleibt die Frage, ob nicht die aktive Tätigkeit lebender Zellen das wirksame Mittel bei der Säftebewegung sein könnte. Diese physiologische Theorie erhielt jedoch einen schweren Schlag durch gewisse, allerdings unzureichende Versuche von Strasburger, welche ihn zum Schlüsse führten, daß das Saftsteigen durch die Einwirkung von Giften, die alles lebende Gewebe notwendig töten müssen, nicht gestört wird. Meine im folgenden zu beschreibenden Versuche führten mich zu Schlüssen, die denen Strasburgers gerade entgegengesetzt sind.
Die folgenden Experimente über die Wirkung stimulierender und giftiger Stoffe zeigen in ganz entscheidender Weise, daß die Saftbewegung im Wesentlichen auf der aktiven Leistung lebender Zellen beruht. Ein abgeschnittener Stengel von Chrysanthemum coronarium wurde dem Eintrocknen überlassen. Die Pflanze hängt über, die Blätter schrumpfen und werden runzelig; die Pflanze sieht wie tot aus. Doch die Zufuhr von Wasser, das Spuren eines stimu lierenden Stoffes enthält, ruft eine wunderbare Veränderung hervor; ein lebhaftes Saftsteigen setzt ein, wodurch der ursprüngliche Turgor wieder hergestellt wird. Der gekrümmte Stengel richtet sich auf und die Blätter breiten sich in ursprünglicher Frische aus, wie man es in der in Fig. 68 wiedergegebenen Photographie sieht. Vollständige Erholung trat in diesem Fall schon nach einer Zeit von 15 Minuten ein. In scharfem Kontrast hierzu stand der Parällelversuch, worin dem welken Stengel giftige Formaldehydlösung zugeführt wurde. Die betreffende Pflanze erholte sich nicht, sondern erfuhr eine gänzliche Zersetzung zu. einer verschwommenen Masse sterbenden Gewebes (Fig. 69).
 |
 |
| Fig. 68. Vollkommene Wiederaufrichtung eines abgeschnittenen Sprosses mit welken Blättern nach Zufuhr von Wasser durch die Schnittfläche (Chrysanthemum). Die Photographien zeigen die Pflanze vor (rechts) und nach der Wasserzufuhr (links). |
Fig. 69. Photographien eines welken, abgeschnittenen Sprosses von Chrysanthemum; er steht in einer Formaldehydlösung, welche verstärktes Welken anstatt der Wiederaufrichtung durch Wasseraufnahme wie in Fig. 68 bewirkt. |
In der nächsten Versuchsreihe brachte ich gewisse Modifikationen an; auch verwendete ich eine andere Spezies. Ein aufrechter Stengel von Centaurea wurde in ein Gefäß P eingestellt, welches eine giftige Lösung von Kaliumcyanid enthielt; ein welker Stengel derselben Pflanze wurde gleichzeitig in das Gefäß S eingestellt, das eine stimulierende Lösung enthielt. Die entgegengesetzte Wirkung der giftigen und der stimulierenden Lösung tritt aufs klarste in den beiden Photographien rechts hervor (Fig. 70). Das vorher aufrechte Exemplar wird unter der Wirkung des Giftes vollständig welk, während der welke Stengel durch das Stimulans ein gesundes, kräftiges Aussehen erlangt.
 |
| Fig. 70. Frischer Stengel, in eine giftige Lösung, und welker Stengel, in eine stimulierende Lösung eingestellt (links). In der giftigen Lösung wird der Stengel welk und stirbt, während der vorher welke Stengel sich wieder aufrichtet (rechts). |
Die hiedurch gewonnene Erkenntnis beweist zur Genüge, daß die Fortbewegung des Saftes wesentlich auf der aktiven Tätigkeit lebender Gewebe beruht. Aber die unbestimmte Annahme einer Lebenstätigkeit gibt noch keine vollständige Erklärung der Erscheinung. Es ist notwendig, weiter das Wesen der zugrundeliegenden Kräfte zu bestimmen, zu ermitteln, in welcher Weise diese Kräfte angreifen und durch was für Mittel ein bestimmt gerichteter Säftetransport zustandekommt.
Um zu einer vollständigen Lösung des Problems zu gelangen, mußte ich diverse Versuchsmethoden ersinnen und verschiedene Apparate konstruieren. Die Resultate, die ich erhielt, führten zu dem Schluß, daß der Saft in den Pflanzen durch einen Mechanismus bewegt wird, der im wesentlichen demjenigen ähnlich ist, welcher die Zirkulation des Blutes beim Tier im Gang erhält.
BEWEGUNG DES BLUTES BEIM TIER
Der Blutkreislauf kommt beim höheren Tier durch die selbsttätigen, ununterbrochenen Pulsationen eines kontrak-tilen Organs zustande, welches wir als Herz bezeichnen. Wir müssen uns freilich von vornherein einen verallgemeinerten Begriff vom Herzen und von seinem Mechanismus bilden, denn es wäre ungereimt, in der Pflanze nach einem so hoch entwickelten komplizierten Organ zu suchen, wie es z. B. das menschliche Herz ist. Vergleichbar mit der Pflanze wäre eher etwa das Herz der niederen Tiere; so ist bei Amphioxus das Herz ein verlängertes, röhrenartiges Organ, und die in ihm enthaltene Nährflüssigkeit wird durch peristaltische Kontraktionswellen, welche fortschreiten, weiterbewegt. Auch bei den höheren Tieren ist das Herz im Embryonalzustand eine verlängerte Röhre. Das wesentliche Merkmal des Herzmuskels beim Tier ist die rhythmische Pulsationsbewegung, deren Geschwindigkeit durch bestimmte Außeneinflüsse in bestimmter Weise verändert wird.
So schlägt das Herz unter der Wirkung bestimmter stimulierender Stoffe schneller und pumpt das Blut rascher vorwärts, was einen schnelleren Umlauf zur Folge hat. Ein Stoff, der einen Depressionszustand herbeiführt, hat genau die entgegengesetzte Wirkung. Ferner ist ein Innendruck von gewisser Höhe notwendig, damit die Pulsation des Herzens beginnen kann. So läßt sich das ruhende Herz der Schnecke zum Pulsieren bringen, wenn der Innendruck künstlich erhöht wird. Die Tätigkeit des Herzens wird innerhalb gewisser Grenzen durch Temperaturerhöhung angeregt und durch die Einwirkung von Kälte vermindert. Eine kleine Dosis eines Narkotikums, wie Äther, beschleunigt die Herztätigkeit. Ein starkes Narkotikum wie Chloroform bewirkt am Beginn eine unmittelbare Stimulation, auf welche eine Depression und bei fortgesetzter Ein-. Wirkung ein völliger Stillstand folgt.
Ich will nun zu zeigen versuchen, daß jeder Faktor, der die Tätigkeit des Herzens erhöht und so das Tempo des Blutumlaufes beschleunigt, auch die Säftebewegung in der Pflanze beschleunigt; und daß umgekehrt jedes zum Depressionszustand führende Mittel den Säftefluß hier wie dort verlangsamt. Die Schwierigkeit, die dabei zu überwinden war, lag darin, daß vordem keine befriedigende Methode zur Verfügung stand, die normale Geschwindigkeit des Saftsteigens und deren Veränderung beim Wechsel der Außenfaktoren messend zu untersuchen.
DAS BLATT ALS INDIKATOR DER SAFTBEWEGUNG
Es gelang mir diese Schwierigkeiten hinwegzuschaffen, indem ich das Blatt als Indikator für die Saftbewegung benützte. In einer eingetopften Pflanze bewirkt Trockenheit früher oder später ein Welken der Blätter, nach dem Begießen richten diese sich wieder auf. Hier bewirkt nun jedes Mittel, das die Lebhaftigkeit des Saftsteigens erhöht-, auch eine rasche Aufrichtung des Blattes; andererseits führt jede Verminderung der Saftsteigetätigkeit durch ein Depressionsmittel eine Senkung des Blattes herbei. Die Bewegungen sind aber zu langsam, um leicht beobachtet werden zu können; sie werden indes deutlich, wenn man sie stark vergrößert, wie das die folgende Vorrichtung erlaubt.
DER ELEKTRISCHE PHYTOGRAPH
Dies ist ein sehr feiner Apparat, welcher der Beobachtung der wechselnden Geschwindigkeit des Saftsteigens dient. Das Blatt wird mit Hilfe eines dünnen Seidenfadens an dem Zeichenhebel befestigt, welcher die Bewegungen des Blattes auf das zehn- bis hundertfache vergrößert. Die Registrierkurve wird auf einer berußten Glasplatte verzeichnet. Der Hebel zeichnet vermittels einer elektromagnetischen Vorrichtung eine Reihe von Punkten auf der sich bewegenden Platte in Abständen von zwei bis zu zehn Sekunden auf (Fig. 71). Die Reaktionskurve, welche das Blatt eines abgeschnittenen Sprosses gibt, ist praktisch dieselbe, wie diejenige einer intakten Pflanze mit Wurzeln; ein abgeschnittener Sproß läßt sich aber leichter und bequemer handhaben.
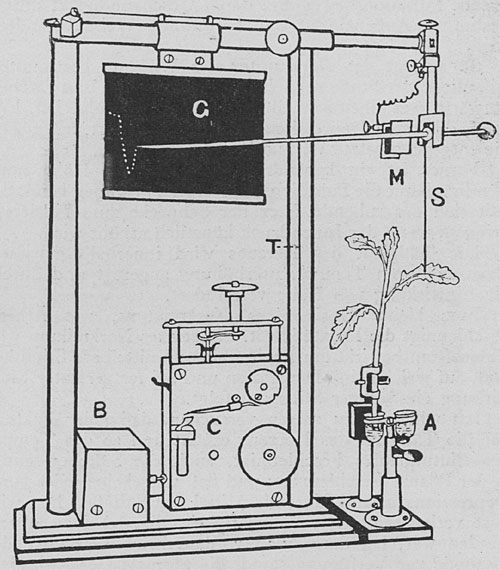 |
| Fig. 71. Der elektrische Phytograph. |
Zwei kleine Probegläschen mit verschiedenen Lösungen werden an einem drehbaren Stativ befestigt, das sich mit Hilfe einer Handschraube heben und senken läßt. Das Schnittende des Sprosses kann so in rascher Folge mit verschiedenen chemischen Agentien, mit Depressions- und Stimulationsmitteln oder mit kaltem und warmem Wasser behandelt werden.
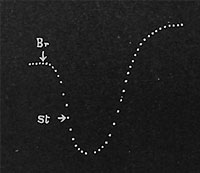 |
| Fig. 72. Aufeinanderfolgende Hemmung und Beschleunigung des Saftaufstieges durch elektrische Depressions- und Stimulationsmittel. |
Um die Ähnlichkeit in der Wirkung des Bewegungsmechanismus bei der Pflanze und beim Tier zu demonstrieren, will ich eine ausführliche Beschreibung der entgegengesetzten Effekte geben, welche Depressions- und Stimulationsmittel bewirken. Das Blatt befand sich in horizontaler Gleichgewichtslage. Zufuhr von Kaliumbromid-lösung brachte eine so starke Depression, daß das Blatt eine rasche Fallbewegung erkennen ließ. Die Zufuhr einer kleinen Dosis Kampfer, welcher ein Stimulationsmittel ist, hatte das Aufhören der Senkung und sodann eine Hebung des Blattes zur Folge (Fig. 72). Man darf daraus schließen, daß der Säftenachschub zum Blatt in dem einen Falle vermindert, im anderen erhöht worden war und daß die angewandten Reagentien lebende Zellen beeinflußt haben müssen, durch deren Tätigkeit der Wassernachschub zustande kommt.
 |
| Fig. 73. Der Effekt abwechselnder Einwirkung von Kälte und Wärme; in der Kälte (C) Hemmung des Saftsteigens und Senkung der Kurve, in der Wärme (H) Beschleunigung des Saftsteigens und Anstieg der Kurve. |
Die Wirkung der Trockenheit zeigte sich in einem Fallen des Blattes. Hier hatte die Beförderung des Saftes eine Unterbrechung erfahren, wahrscheinlich infolge der Verminderung des hydrostatischen Innendruckes der aktiven Zellen. Der Effekt abwechselnder Einwirkung von kaltem und warmem Wasser bestand in aufeinanderfolgender Verminderung und Erhöhung der Geschwindigkeit des Saftaufstieges, wie dies die Kurve in Fig. 73 klar erkennen läßt.
Verdünnter Äther ergab eine Beschleunigung des Saftsteigens und eine rasche Aufrichtung des Blattes. Eine Stimulierung mit folgender Depression zeigte sich bei der Wirkung von Chloroform, sie war an der anfänglichen Aufrichtung des Blattes und der folgenden raschen Fallbewegung kenntlich.
Einige der wichtigsten Ergebnisse dieses Kapitels mögen nun zusammengefaßt werden. Die Tatsache, daß in einer mit Gift behandelten Pflanze die Saftbewegung stillsteht und dauernd vernichtet wird, beweist, daß diese auf der aktiven Tätigkeit lebender Gewebe beruht. Der Stillstand und die Wiedererneuerung des Saftsteigens unter dem Einfluß von Kälte und Wärme und die charakteristische Beschleunigung und Verlangsamung unter dem Einfluß von Mitteln, die die Herztätigkeit stimulieren oder herabsetzen, weisen weiter darauf hin, daß der Mechanismus der Säftebewegung bei der Pflanze und beim Tier grundsätzlich ähnlich ist. Rein physikalische Mechanismen können jenen charakteristischen Reaktionen in keiner Weise zugrunde liegen.
Erläuterung der Fußnoten.
4. Noll in Strasburger-Noll-Schenk-Schimper, Lehrbuch der Botanik f. Hochschulen, 5. Aufl., 1902, S. 155.




